個体数情報は少ないが10,000羽程度と推定されている(Chia 2011)。この結果は約6,700羽の成鳥個体に相当している。減少理由は、生息地の破壊のだと考えられており(Ferguson-Lees and Christie 2001)、原生林の伐採が脅威となっている(Ferguson-Lees and Christie 2001)。インドのミゾラム州での狩猟も、潜在的な脅威として確認されている(Ved and John 2011)。インドのケララ州では、カンムリワシ(Crested Serpent Eagle)が本種をを捕食・攻撃しているのが観察されている(Vishnudas 2007)。
Source: BirdLife International (2025) Data Zone. Retrieved from http://datazone.birdlife.org
<分類>
5亜種に分類される。
〇 A. j. jerdoni
インド北部からミャンマー、インドシナ、スマトラ島に分布する。
〇 A. j. ceylonensis
インド南西部からスリランカに分布する。他亜種に比べて体が小さい。上面や胸部が淡色で、バンドが細目で色が薄い。尾羽の鷹斑も幅が狭い傾向にある。
〇 A. j. borneensis
ボルネオ島に分布する。
〇 A.j.magnirostris
フィリピンのミンダナオ島、パラワン島、サマール島に分布する。
〇 A. j. celebensis
インドネシアのスラウェシ島、トギアン諸島、バンガイ諸島、スラ諸島に分布する。
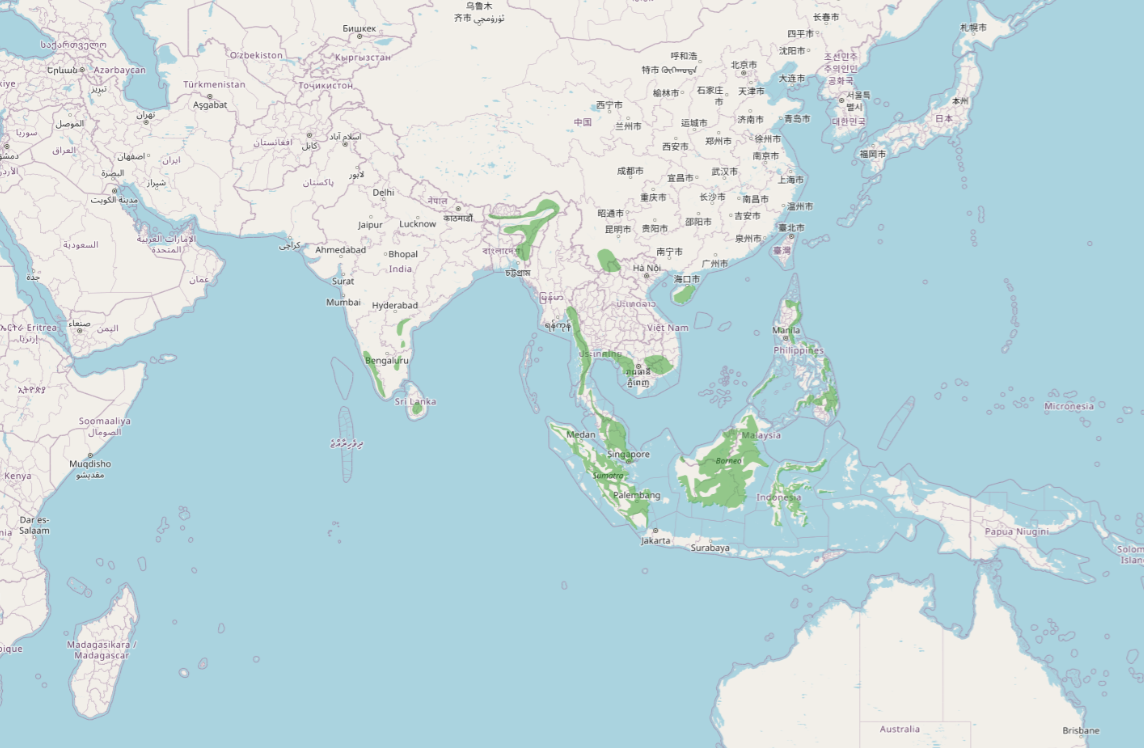
マレー半島、インドネシア、マレーシア、台湾、中国南東部、ベトナム、カンボジア、タイ、スリランカ、インド、ネパールなどの山岳地に分布する。東南アジアの森林地帯に分布域を有する。個体数の減少傾向は続いているようだが、その減少は「絶滅危惧 II 類」に達するほど急速ではないとしている。これらの理由から、この種は「LC(低懸念)」と評価されている。
世界全体の個体数は約 1万~10万羽と推定されている(Ferguson-Lees et al. 2001)。一方、中国国内の繁殖ペア数は 100ペア未満で、台湾では100~10,000 ペアと推定されている(Brazil 2009)。森林分布に大きく依存しており、過去3世代の間に、その生息域内の樹木被覆率は 18.3% 減少したと推定されている(Global Forest Watch 2022、Hansen et al. 2013)。従って、この森林被覆率の低下により、同期間にこの種の個体数が 1~19% 減少した可能性があると推測され、その減少率は 15~19% と推定されている。
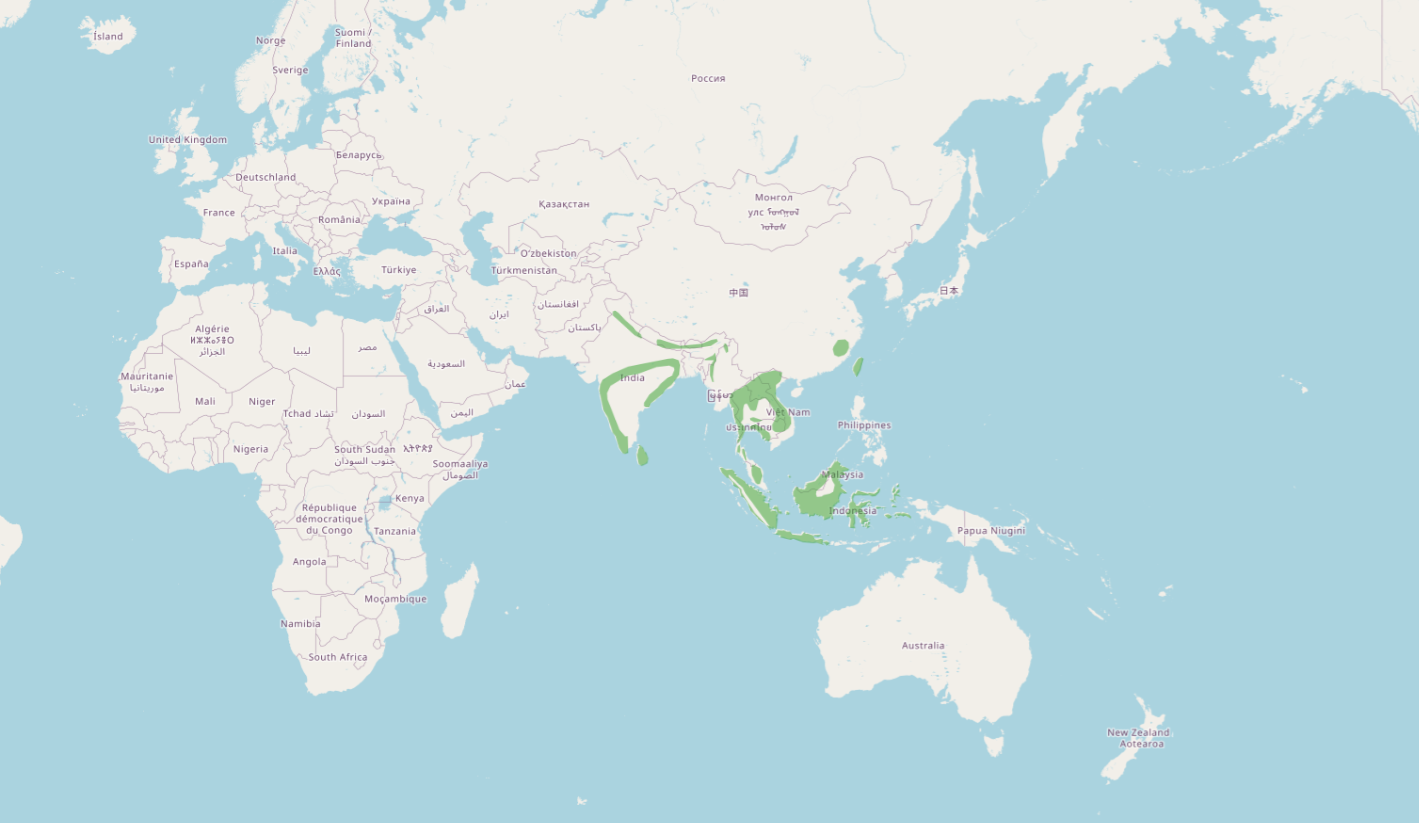
カザノワシの分布図 Source: BirdLife International (2025) Data Zone. Retrieved from http://datazone.birdlife.org
世界全体の個体数は成鳥20万羽と推定されている(Partners in Flight 2020)。北米大陸(特にアメリカとカナダ)における総個体数は成鳥20万羽と推定している(Partners in Flight 2020)。北米大陸(アメリカとカナダ)における本種の個体数は、わずかに増加傾向にあると推定されており、これは3世代にわたる個体数の急速な増加に相当する(Partners in Flight 2024)。この地域が本種の世界分布域の大部分を占めるため、世界全体の個体数は3世代にわたって急速に増加していると見なされている。北米では40年間で779%増加、10年当たり72.2%増加に相当すると評価している(Butcher and Niven 2007)。
1960年代には約30ペアと、過去最低水準にまで減少したが、1970年には38ペアに回復し、2004年までに約200ペアに増加した(González and Oria 2004)。その後も個体数は大幅に増加したが、近年は限界値に達したとみられている(S. Cabezas-Díaz in litt. 2020)。2020年時点で、世界全体の繁殖ペア数は530~540に達したと推定されている(BirdLife International in prep.)。ポルトガルでは20年以上繁殖が確認されていなかったが、2003年に再び定着が確認されて以降、徐々に個体数が増加しており、2016年には16ペア、2019年には17ペアの繁殖が確認された(S. Cabezas-Díaz and J. C. Atienza in litt. 2013;S. Cabezas-Díaz in litt. 2020)。
この個体数回復の主な要因としては、送電線や毒物による死亡率の低下を目指した保全措置が挙げられる。これには補食、巣の修復、再導入、繁殖期の擾乱の減少などが含まれる(González et al. 2007, López-López et al. 2011, J. R. Garrido in litt. 2013, S. Cabezas-Díaz and J. C. Atienza in litt. 2016, M. Ferrer in litt. 2016, J. R. Garrido in litt. 2017)。一方で、近年は送電線、射撃、風力発電機との衝突による死亡率が上昇しており、毒殺も依然として深刻な脅威である(S. Cabezas-Díaz in litt. 2020)。
イベリア半島での繁殖状況の改善に伴い、20世紀前半に繁殖種として絶滅したモロッコでは、分散個体の記録が増加している(Thevenot et al. 2003, Maghreb Ornitho 2017)。スペインのカディスに再導入された個体群では、衛星タグを装着した6羽の若鳥のうち複数がモロッコへの渡りを行ったことが確認されている(Morandini et al. 2016)。また、未成熟個体がアルジェリア、モーリタニア、西サハラへ移動した事例もある(Morandini et al. 2020)。モロッコでの繁殖群復活の可能性もあるが、同国では感電による死亡率が極めて高いことが報告されている(Godino et al. 2016)。
種全体の個体数は、530~540の繁殖ペア(成鳥1,060~1,080羽)と推定され、若鳥を含めた総個体数は約1,600羽とされている(BirdLife International 準備中)。本種の個体群は非常に小さく、毒殺、感電、射殺、風力発電機との衝突、餌資源の不足など、多くの脅威に対して集中的かつ継続的な管理措置に依存しており、「VU(危急)」に分類されている。20世紀末から21世紀初頭にかけては個体数が着実に増加したが、現在は停滞期にあるとみられている。
1989年~2004年の16年間に記録された267件の非自然死の分析では、年間平均15.1個体が死亡していた。その主な死因は感電(47.7%)および中毒(30.7%)である(González et al. 2007)。死亡事例の約40%は狩猟や家畜保護活動に関連していたが、ほとんどは偶発的な事故だった(同上)。2015年から2016年には毒物による死亡が15件報告されており、特に懸念される問題となっている(S. Cabezas-Díaz and J. C. Atienza in litt. 2016)。1990年代後半の個体数減少は、毒餌による中毒死が主因とされ、ドニャーナ国立公園の個体群は深刻な影響を受けた(Ortega et al. 2009, Ferrer et al. 2013)。
感電死は依然として重大な問題であり、緩和策により死亡数は大きく減少したものの(López-López et al. 2011)、特に若鳥において依然深刻である。2015年には、モロッコへ渡った6羽の衛星タグ付き個体のうち1羽が感電死し、その後新たに設置された高圧送電線付近でさらに4羽の若鳥の死体が発見された(Godino et al. 2016)。モロッコに渡った個体数は少数と見られているが、送電線が繁殖群の再定着を阻害する深刻な要因となる可能性がある。
スペイン南部のアンダルシアでの緩和措置は概ね成功しているが、2010年以降でも11羽の感電死が確認されている(J. R. Garrido in litt. 2016)。過去10年間で、スペインおよびポルトガルで複数の個体が射殺されたことも報告されており(S. Cabezas-Díaz and J. C. Atienza in litt. 2016, J. R. Garrido in litt. 2016)、狩猟が依然として脅威であることが示唆されている。加えて、獲物の肉に含まれる鉛弾の摂取が、特定地域で問題となる可能性もある(González and Oria 2004, Pain et al. 2005)。中毒や射殺の脅威は、繁殖域全体において土地所有者との連携を継続することの重要性を示している。
スペインでは1973年から2002年にかけて、主な獲物であるウサギの個体数が55%減少した(Virgós et al. 2007)。ウイルス性出血性疾患の流行や、狩猟地の管理方法の変化が獲物の可用性低下の要因となっている(Cabezas-Díaz et al. 2011)。一部の地域では、灌漑を伴う農業の集約化や都市開発の拡大が生息地の適性を低下させ、将来の個体数の増加を制限する可能性がある。また、これらの地域では人間との衝突の増加も報告されている(J. R. Garrido in litt. 2016)。
再生可能エネルギー開発による広大な土地利用は、現時点では明確な負の影響は確認されていないが、2012年にはアンダルシアで1羽が死亡した事例がある(J. R. Garrido in litt. 2016)。この脅威は、今後風力発電所の設置が増加する中で拡大する可能性がある(S. Cabezas-Díaz in litt. 2020)。また、繁殖中の巣の周囲での人間活動は、抱卵中の親鳥を擾乱し、孵化率の低下を引き起こす可能性がある(González et al. 2006, Margalida et al. 2007)。加えて、送電線の安全対策など鳥類保護に必要なモニタリングと緩和措置の実施には、資金不足が大きな制約となる恐れがある(S. Cabezas-Díaz and J. C. Atienza in litt. 2016)。
イギリス諸島、スカンジナビア半島、ロシア北部、シベリア、南は地中海、コーカサス、ヒマラヤ、日本を含むヨーロッパと北アジアに広く分布している。また、アラスカ西部および中央部、カナダ、アメリカ、南はメキシコまで、北アメリカにも広く生息している。生息域の大部分では留鳥性だが、北米、スカンジナビア、ロシアの最北の個体群では部分的に渡り性である(Squires et al.)
日本では沖縄県を除く全土に分布し、繁殖が確認されている。沖縄県では越冬期に確認されることがある。北海道など豪雪地帯のものは南下する。
ヨーロッパの個体数は234,000~380,000羽と推定されている(BirdLife International In prep.) ヨーロッパは世界的な生息域の約26%を形成しているため、世界的な個体数規模を推定すると90万羽~146万羽になるが、この推定値については検証が必要である。カナダとアメリカにおける個体数は成鳥個体数で21万羽と推定されている(Partners in Flight 2020) これらの情報から世界中の推定成鳥個体数は100万羽~250万羽と推定される。
19~20世紀におけるヨーロッパでの著しい減少は、ハンディングと森林伐採によるもので、1950~1960年代における減少は、農薬や重金属による中毒の結果であると考えられている。ノルウェーにおけるオオタカの巣の研究によると、残留性有機汚染物質が血液、羽毛に存在していることが示唆されている(Briels et al.) 鷹狩りのための巣の強奪と同様に、迫害も脅威であり続けている(Orta and Marks 2014)。また、風力発電所開発の影響に対しても非常に脆弱である(Strix 2012)。木材伐採は営巣個体群にとっての主要な脅威であり、巣や巣台は定期的に破壊され、また伐採作業によって破壊されるが、繁殖成功への影響は不明である(Squires et al.) 巣の近くでの伐採活動による撹乱も巣の失敗の原因となる(Boal and Mannan 1994)。北米西部では山火事の頻度と強度が増加し、ねぐらや採餌に適した場所が減少している(Blakey et al.) ヨーロッパのいくつかの国々における減少は、農業と林業の激化に伴う食糧供給の 減少、およびワシミミズクBubo buboとの巣場競争の激化に関連している(Rutz他 2006年)。世界的には個体数の変化は不明であると考えられている。この種は北米では過去40年間にわたり安定した個体数傾向を示している (Breeding Bird Survey、Christmas Bird Countのデータ: Butcher and Niven 2007)。ヨーロッパでは、個体数は24年間(3世代)で59%減少していると推定されている。最近、オランダ東部(Rutz他、2006年)、ドイツ(Gedeon他、2015年)、デンマーク(2005~2014年、年率-8%、Nyegaard他、2015年)、フィンランド(Valkama他、2011年)で減少が記録されている。
この種は極めて広い生息域を持つため、生息域の広さ基準では、絶滅危惧Ⅱ類の閾値に近づかない。個体数が減少傾向にあるように見えるにもかかわらず、その減少は個体数傾向基準のもとで、絶滅危惧Ⅱ類のしきい値に近づくほど急激なものではないと考えられる。個体群サイズが非常に大きいため、個体群サイズ基準における絶滅危惧Ⅱ類のしきい値に近づかない。これらの理由からこの種は「低懸念(LC)」と評価される。総個体数を100,000個体、成熟個体数を67,000個体と推定している(Ferguson-Lees and Christie 2001)。国内の個体数は中国で約10,000~100,000繁殖ペア、台湾で約10,000~100,000繁殖ペアと推定されている(Brazil 2009)。他の地域からの情報がないが、成熟個体数40,000~400,000の範囲に位置づけられる。森林の減少が続いていることから個体数の減少が疑われている(Ferguson-Lees and Christie 2001)。2001年から2020年の間にこの種の生息域全体で、8.9%の森林被覆が失われ(Grobal Forest Watch 2021)、 2016~2020年の間に森林被覆の3.3%が失われ(Grobal Forest Watch 2021)、3世代にわたって予測すると10.3%に相当する。この種は森林の生息環境に大きく依存している(Ferguson-Lees and Christie 2001)。現在、この種に対する他の脅威は知られていないため、森林の消失と同様の割合で減少していると考えられる。インド、フィリピン、ジャワ島の低地における森林伐採がこの種にとっての主な脅威であるとされる(Clark and Marks 2014)。
極東にのみ分布する。ロシアのカムチャッカ半島、オホーツク海周辺の沿岸地域、アムール川下流域(ゴリン川以南)、サハリン北部およびシャンタルで繁殖する。冬季になると南下し、サハリン、北海道、東北を主な越冬地としている。日本のその他の地域では数が少ない。中国北東部、北朝鮮、韓国でもごく稀に観察される。1991年から2009年にかけて、マガダン地方の内陸河川個体群は繁殖成功率が低下しており、沿岸個体群は同じ期間に繁殖成功率が緩やかに上昇していることから、これらの個体群はそれぞれシンク個体群(減少もしくは絶滅しそうな個体群)とソース個体群(安定的で供給源としての個体群)と考えられている(Potapov et al.) 総個体数は5,000~7,000羽、北方四島を含む日本国内の越冬数は 2,000~2,500羽と推定されている(中川、2009b、2013)。M. McGrady et al. (in litt. 2012)は、世界の個体数を約4,600~5,100個体、うち繁殖ペアは約1,830~1,900組、成熟個体数は約3,600~3,800頭と推定している。しかし、Masterov and Romanov (2014)は、世界の個体数を6,000-7,000と推定しており、これは成熟個体数4,000-4,670にほぼ相当する。しかしながら、近年の観察において8000羽を超えるオオワシが宗谷岬から渡ってきており、総個体数としてはもっと多い可能性が指摘される。
1991~1998年にオホーツク海北部の海岸線および河川沿いで行われた調査では、繁殖に参加したペアの割合とテリトリーペア1組当たりに羽化したヒナの数が著しく減少(70%から35%へ)していることがわかった(Potapov 2000)。最近の同じ地域での調査でも、成功したペア1組あたりに羽化したヒナの数が過去10年間で減少していることが判明している(Potapov et al.) 河川沿いに営巣するワシの繁殖率が低下しているのは、春の洪水の頻度と強度が増加していることと関連しており、この洪水は狩りの成功を妨げ、その結果繁殖数が減少している(Potapov et al.)。 沿岸のペアにおける繁殖率の減少は、春の氷被覆の減少と関連している(Potapov et al.)。 ロシアでは水力発電プロジェクト、石油化学産業のための大規模な沿岸・ 沖合開発計画、木材の伐採などの開発中に生息地が改変され、絶滅の危機に瀕しているとしている(Ferguson-Lees and Christie 2001)。河川の工業汚染や高濃度のDDT/DDE、PCB、重金属は脅威であると考えられている(Iwata et al.)。 乱獲はロシアと日本で魚類資源の減少を引き起こし、その結果、北海道の鳥類が内陸に移動し、狩猟者が残したニホンジカの死骸をあさる傾向が強まり鉛弾を摂取することによって鉛中毒のリスクにさらされるようになった(Ferguson-Lees & Christie 2001)。日本では2004年に大型動物の狩猟に鉛弾の使用を禁止する法律が導入され、2014年にはさらに鉛弾の所持を禁止する法律が施行されたが、鉛中毒は依然として重大な脅威である(Ishii et al.)。
<分類>
亜種は分類されていないが、チョウセンオオワシH. p. nigerが朝鮮半島で繁殖していた個体群とされていたが、現在は絶滅している。チョウセンオオワシは肩の白色部が暗色でああることが標本から分かっている。
<渡り>
オオワシは集団で渡ることが近年解明されてきており、サハリンやカムチャッカ半島で集結した渡りが見られる。日本においては宗谷岬やオホーツク沿岸でも観察される(環境省自然環境局、2013b,Ueta et al., 2000; McGrady et al.,2003)。1998年の秋の観察ではサハリンから宗谷岬に590羽、一日最大230羽がが観察された(伊藤、1991)。1999-2000年の調査では1カ月に2000羽近く観察され1日最大500羽が観察された(植田ほか,2004)。近年では1日最大1500羽を超える観察例もある。
ヨーロッパでは、繁殖個体数は10,400~14,600つがい、成熟個体数は20,900~29,200羽と推定されている(BirdLife International in prep.) ヨーロッパは世界的な生息域の50~74%を形成しているため、世界的な個体数の推定値は28,200~58,400羽となるが、この推定値についてはさらなる検証が必要である。
巣の保護、安全な餌の提供、バイエルンなど特定の地域への再導入などの保護対策により、個体数は局所的に増加している(Ferguson-Lees and Christie 2001)。ヨーロッパの個体数は純粋に増加している(BirdLife International in prep.)。ロシアの一部(Shukov 2019)と日本でも個体数が増加している(Shiraki 2018)。この種は生息域が非常に広いことや個体数が増加傾向であることなどから、この種は「低懸念(Least Concern)」と評価される。
この種に与える脅威は湿地帯の損失と劣化、人間による撹乱と迫害、環境汚染、風力発電機との衝突(Krone and Scharnweber 2003)、毒物の無差別使用などがある。近代的な林業手法により、営巣に適した生息地が減少している(Orta et al.) 有機塩素系農薬や重金属による汚染は、特にバルト海沿岸地域で繁殖成功率の低下をもたらした(Orta et al.) アジアやロシアでは伐採や石油産業開発の増加により若干の減少が見られるが、ヨーロッパでの増加がそれを上回っている。フィンランドでは、使用済み弾薬による鉛中毒事故が依然としてかなりの死亡原因となっており 全体として、人為的要因がワシの死亡原因の60%を占めていることが明らかになった(Isomursu et al)。大木が失われたことで営巣に適した場所が不足し、人間への馴化が進んだことで、ワシは都市に近い場所で営巣するようになった(Shiraki 2018)。このためワシと交通の衝突事故が増加し、繁殖成功率の低下につながっている可能性がある(同書)。風力タービンとの衝突はさらなる死亡要因であるが(Heuck他、2019;Matsuda他、2021)、そのリスクは順応的管理によって軽減することができる(Matsuda他、2021)。鳥類は時折、鳥インフルエンザウイルスの発生によって脅かされる(Krone et al.)日本でも橋梁や道路などにオジロワシが衝突しないようにポールや壁などが設置される試みが行われている。
<分布など>
オーストラリアおよびパプアニューギニアの一部に生息する。この種は生息域が非常に広いため、生息域の広さの基準では絶滅危惧Ⅱ類(Vulnerable)の閾値に達しない。7,680,000km²の範囲に推定10万羽が生息している。個体数は増加傾向にあるとされることが多いが、射殺、捕獲、意図的な毒殺、生息地の破壊などにより個体数は減少していると考えている研究者の方が優勢である。これらの理由から、この種は「低懸念(Least Concern)」と評価される。腐肉を好み、交通事故死したカンガルーやワラビー、羊、ダチョウなどを食べる。その為、二次被害で交通事故死する例がある。亜種<i>A. a. fleayi</i>は1999年、環境保護および生物多様性保全法により絶滅危惧種に分類された。彼らの食餌に占める羊の割合は3%以下であり、捕食する羊の99%は腐肉であるという調査結果があるにも関わらず、羊にとっての脅威とみなされている。
<分布など>
ロシア南部、北朝鮮、中国北東部、韓国北部と中国中部南東部で繁殖するが分布が離れている。冬季はフィリピン、マレーシア、インドネシアなどに南下して越冬する。生息域が非常に広い。個体数は減少しているが、現在のところ減少率は個体数傾向基準(10年間または3世代にわたり30%以上減少)の「絶滅危惧(Vulnerable)」のしきい値には達していない。そのため、「低懸念(Least Concern)」と評価される。ただし、個体数減少の割合が増加した場合、この種は絶滅危惧種に指定される可能性がある。そのため、継続的な個体数のモニタリングが必要である。中国、台湾では、気象レーダーを用いて春の渡りの時期に上空を通過する個体数を推定している。2004年には最大41万個体が推定された(Chen et al. 2004)、少なくとも225,935個体がわずか4月5日の間に上空を通過した(Sun et al.)。 Germi他(2009)は、2007年9月20日から11月30日の間に22.5万羽を数え、少なくとも35万羽のアカハラダカがインドネシア東部に渡来すると推定している。これらの合計は総個体数の最良推定値であるため、40万~100万個体が生息すると推定される。
この種の越冬範囲内での土地利用の変化は、ねぐらとなる生息地の喪失を もたらす可能性がある(Germi et al.) 例えば、バリ島での伐採作業は、ねぐらとなる場所の利用可能性を減少させる可能 性がある(Germi 2005)。日本と韓国では、二次林、特に工業地域周辺における営巣資源の損失が報告されている (Choi他、2008年)。航空機によるバードストライクも潜在的な脅威である(Tang et al.) 2015年にはバリ島の市場で1羽のアカハラダカが売りに出されているのが発見され、インドネシアでは19羽がSNS上で売りに出されていたが、取引の規模は大きくないと思われる(Germi and Waluyo 2006; Gunawan et al.)
<分類>
亜種は分けられていない。
<渡り>
長距離の渡りを行う種である。日本、台湾、中国、フィリピン、インドネシアの島々を経由し、個体数の大部分は台湾、中国、インドネシア北部のサンギヘ諸島およびタラウド諸島を通過する(Chen et al.2004, Germi et al.2009, Sun et al.2010)。第二のルートは東南アジア大陸を通り、タイ、マレーシア半島、スンダ列島に沿ったインドネシアへの通過が確認されている(Orta & Kirwan 2020)。近年の研究では渡りで夜間飛行を行うことが知られる。韓国を出発した個体の多くは夜通し飛行し、八重山列島に到着する個体が多い。長崎県を通過した個体は沖縄本島を経由した後、直接ルソン島に到達する個体が多い。群れになりやすく、日本では対馬、長崎県などで大規模な渡りが観察される。奄美大島、沖縄本島、八重山列島でも見られる。台湾南部でも大規模な渡りが観察される。
<分布など>
モンゴル、インド、パキスタンなどの高標高地を中心として繁殖分布している。中東のトルコ、シリア、イラクやヨーロッパの一部、アフリカ東岸にもみられる。絶滅した地域も多い。個体数が急速に減少しているという証拠があるため、「NT(純絶滅危惧)」に指定されている。生息域の一部では保護活動により個体数が安定または増加しているが、ヒマラヤ山脈とインドの一部、および孤立した南アフリカの個体群では急激な減少が疑われている。減少の原因は、ジクロフェナク(家畜の抗炎症や鎮痛薬だがヒゲワシにとって毒性がある)によるものだけでなく、風力発電施設の衝突、撹乱の増加、餌の減少による死亡もある。総個体数を1,000~10,000羽と推定している(Ferguson-Leesら2001)が、ヨーロッパでは630~960ペアと推定されており、個体数としては1890~2880羽が生息しているとしている(BirdLife International in prep.)。 ネパールの個体数は2010年に約500羽と推定されている(K. Paudel and T. Galligan in litt. 2014)。イラクでは20ペア以下(R. Porter in litt. 2013)、アラビア半島では成熟個体数が100羽以下(Symes et al.) エチオピアには数百ペアがいると推定されている(I. Angelov in litt. 2011)。2011年現在、ケニアでは3ヶ所、タンザニアでは6ヶ所以上の巣が確認されているのみで、ウガンダの個体数は不明である。モロッコには6~10ペアが生息していると推定されているが(Cuzin 2019)、アルジェリアでの生息状況に関する最近の情報はなく、チュニジアでは絶滅したと考えられている(F. Cuzin in litt. 2011)。従って、北アフリカの総個体数は8~14ペア程度と推定される。南アフリカでは、個体数は約100ペアと推定されている (S. Krüger in litt. 2012)。したがって、世界全体の推定個体数は2,500~10,000羽となる。
個体数の動向は種の分布域全体で異なっている。ヨーロッパの個体数は1980年以降増加しているが、これは主に再導入プログラムなどの保護活動によるものである(BirdLife International in prep.)。しかし、ネパールのアッパームスタングでの調査では、2002年から2014年の間に個体数が89.3%減少したことが記録されており、ジクロフェナク中毒が原因と疑われている(Paudel et al.2016)。インドのヒマラヤでは、近年減少が認識されている(P. Trivedi in litt. 2013)。かつてはヒマラヤ西部と中部でよく見られたが、近年はヒマラヤ中部下部では攪乱のためかあまり観察されなくなり(R. Naoroji in litt. 2011)、ウッタラーカンドでは1990年代後半から明らかに減少している(M. Sharma in litt. 2014)。ラダックとヒマラヤ山脈の高地に生息する個体群は、安全であると考えられている(R. Naoroji in litt. 2011)。eBirdにおけるインドでのヒゲワシ目撃報告の頻度は、2000年から2018年の間に約60%減少しており、この地域における個体数の減少を示している(SoIB 2020)。カザフスタン南東部では個体数は安定しているようである(S. Sklyarenko in litt. 2011)。イエメンでは1980年代初頭から減少している(R. Porter in litt. 2013)。トルコにおけるこの種の生息域と個体数も近年減少している(K. A. Boyla in litt. 2014, BirdLife International 2015)。アルメニアでは1990年代以降、個体数は安定している(M. Ghasabyan in litt. 2011)。アフリカ南部の孤立した個体群では、死亡率の増加(Kruger et al. 2015)と繁殖成功率の低下(Kruger & Amar 2017)により、1980年代初頭から繁殖域が約27%減少し、繁殖テリトリー数は1960~1999年と2000~2012年の間に32~51%減少している(Krüger et al. 2014, S. Krüger in litt.) 全体として、個体数は過去3世代で20~29%減少した疑いがある。
ヨーロッパでは、飼育下繁殖と再導入プログラムがオーストリア、フランス、イタリア、スイスのアルプスで実施され、個体はその後フランスの他の地域にも広がった(Snow and Perrins 1998, Frey and Walter 1989)。スペインの一部では再導入計画が進行中である(J. A. Gil Gallus in litt. 2011)。ピレネー山脈では給餌場が設置され、その結果この種の個体数が増加しており、 この種の生息域全域に同様の給餌場が設置されれば、世界的な個体数密度が向上する可能性が ある(Ferguson-Lees and Christie 2001)。しかし、これらは個体数の増加や個体の生存率の向上に役立っている一方で、ハゲワシに悪影響を及ぼす可能性がある。例えば、生息地の飽和につながり、これらの地域では個体のテリトリーが重複し、生産性の低下につながる可能性がある(Carrete et al.)。
ロシア、中国北東部、ウクライナや北欧などで繁殖し、冬季は南下する。日本、韓国、台湾、中国からヨーロッパまで広域に広がる。モンゴルやアフリカ、インドではほとんど見られない。この種は生息域が非常に広い。ヨーロッパ個体群の繁殖雌数は56,300~86,600羽と推定されており、112,000~174,000羽の成鳥が生息すると推定されている(BirdLife International in prep.)。 ヨーロッパは全世界の生息域の約34%を占めていることから、全世界の個体数は330,000~512,000羽の成鳥が生息していると推定される。この推定は疑義が残る。生息域の改変が個体群サイズに及ぼす影響については不確実であるため、個体群の傾向 を決定することは困難である。ヨーロッパにおける全体的な傾向は不明確であるが(BirdLife International in prep.)、いくつかのヨーロッパ諸国で減少が報告されていることから、減少していることが疑われる(Fernandez-Bellon et al.)。 ヨーロッパでは生息域がここ数十年で縮小している(Keller et al.)。 アジアのロシアにおける傾向は不明である。この種は減少の危険が少ない、または判断が困難とされており、「低危険種(LC)」と評価されている。
現在の主な脅威は、農業の激化、湿地の消滅、再植林による生息地の変化である。植生の焼却は、繁殖地、非繁殖地ともにアイルランドで最もよく記録される圧力のひとつである(Caravaggi et al.)。 スコットランドのムーアと呼ばれる湿原などは、地域的に依然として深刻な圧力を受けている。300つがい以上の生息地があると推定されているにも関わらず、2013年にはイングランドで1組も営巣に成功していない(Pitches 2013)。2015年から2019年にかけて英国で衛星タグが付けられた117羽のハイイロチュウヒのヒナのうち、2020年6月に生存していたのはわずか15羽で、ほとんどが不審な状況で行方不明になっている(RSPB 2020)。中央ヨーロッパと東ヨーロッパでは違法に射殺されている(Tucker and Heath 1994)。
個体数は狩り、繁殖地および採食地の喪失と劣化により、地域的に減少 していると考えられる(Ferguson-Lees and Christie 2001)。日本でも減少が記録されており、1970年代には28の調査単位で確認されたが、1990年代には1つも確認されなかった(植田他、2006年)。これらの減少は、昔ながらの水田の放棄や開発による生息地の喪失(Ueta et al. 2006)や、圃場整備などによる餌の減少(Fujita et al. 2015)と関連しているとしている。ロシアの個体群も移動ルート沿いの迫害により著しく減少している可能性がある(Orta & Marks 2020)。中国東北部における個体数の減少は、木材生産のための森林管理や農地の開墾による生息地の喪失と関連しているらしい(Deng et al. 2003)。この種は生息域が非常に広いが、生息域の広さ基準では、絶滅危惧Ⅱ類(Vulnerable)の閾値に近づいていない。個体数が減少傾向にあるように見えるにもかかわらず、その減少は個体数傾向基準(10年間または3世代にわたる30%以上の減少)のもとで、絶滅危惧Ⅱ類(Vulnerable)のしきい値に近づくほど急激なものではないと考えられる。個体群規模は非常に大きいことが疑われている。これらの理由から、この種は「低危険種(LC)」と評価される。中国北東部の森林における繁殖地の喪失は、農業開発と木材伐採によるものである (Deng et al.) 日本では、多くの伝統的な水田が放棄されたり、集約農業や住宅地、工業施設として開発されたりして、採食に適した生息地が減少している(Ueta et al. 2006)。台湾(中国)では毎年1,000羽もの鳥が渡りの際に撃たれている(Orta and Marks 2020)。
<分類>
亜種は分けられていない。
<渡り>
日本では各地で観察されているが、宮古島のカウントが日本の個体数と考えられている。1980年に日本の宮古諸島で53,575羽(DeCandido et al. 2004)、1999年に約32,000羽(Nitani 2000)、2020年で11713羽(宮古野鳥の会)が観察されている。台湾南部の恒春半島では2019年に70442羽、タイのインドシナ半島プロムスリ・ヒルで2003年に14,962羽が観察されている (DeCandido 2004)。
<分布など>
ヨーロッパ南部から中央アジア南部、中東に及び、イラン、ヒマラヤ山脈、モンゴルには狭い範囲で生息している。ナミビアと南アフリカにも繁殖個体群がいる。日本では記録がない。ヨーロッパの個体数は23,330-30,300ペアと推定され、これは46,600-60,500の成鳥に相当する(BirdLife International in prep.)。 北アフリカの個体数は約10,000ペアまたは20,000羽の成鳥と推定される(Garrido et al.)。この種は森林破壊、人間による撹乱、迫害、餌生物種の減少により、地域的に減少している(Ferguson-Lees and Christie 2001)。ヨーロッパと北アフリカでは個体数は増加推定がされている(BirdLife International in prep.)。全体として個体数の傾向は安定していると考えられている(Global Raptor Information Network 2021; Orta et al.)が明確な根拠は少ない。インド亜大陸北部とバレアレス諸島の個体群は主に渡り鳥であり、アフリカ南部とアジア南部で越冬する。
生息域が非常に広いため、生息域の広さ基準(生息域の広さ、生息域の広さ/質、個体数、生息域の広さ/質が減少または変動していること、生息域の数が少ないこと、または分断が激しいことと、出現範囲が20,000km2未満であること)では絶滅危惧Ⅱ類(Vulnerable)の閾値に達しない。個体群規模は中程度から大きいかもしれないが、個体群規模基準(10,000個体未満の成熟個体で、10年間または3世代で10%を超えると推定される継続的減少、または特定の個体群構造)の下での絶滅危惧閾値に近づくとは考えられない。これらの理由から、この種は「低危険種(LC)」と評価される。
この種に影響を与えている脅威は、生息地の劣化、直接的な迫害、人間による撹乱などがある(Ferguson-Lees and Christie 2001, Orta and Boesman 2013)。ウクライナでの減少は森林伐採によるものであり(Orta and Boesman 2013)、ヨーロッパにおいては生息地の消失、草地管理の放棄、農業の集約化、都市化、植林、火災によるものである(2013~2018年の期間、EU加盟国が鳥類指令第12条に基づき報告した圧力と脅威のデータ)。西アフリカでは木材伐採、過放牧、焼畑、農薬が影響している(Thiollay 2007)。越冬地における有機塩素系農薬の蓄積は、この種の繁殖成功に影響を及ぼす可能性がある(Tucker and Heath 1994)。かつてスペイン南東部では、有機塩素汚染が個体数減少の一因となった可能性がある (Martinez-Lopez et al. 2007)。また、潜在的な風力発電開発の影響にも非常に弱い(Strix 2012)。1990~2006年にスペインで発生したブーテッドイーグル死亡例の19.5%は送電線が原因であり、その件数は期間を通じて増加している(Martinez et al.)。 同調査では、違法な人為的による死亡例の32.5%を占めていたが、調査期間中に件数は大幅に減少した。そのほとんどが射殺(19.5%)であったが、捕獲、わな、毒殺も発生していた(Martinez et al.) アルメニアでは密猟も脅威として認識されており、その主な理由は、ニワトリやハトなどの家禽類を殺してしまうのではないかという懸念によるものである(Aghababyan & Stepanyan 2020)。2016年にはマルタ島で渡り中に数羽が射殺された(RSPB 2017)。モロッコの市場でのこの種の取引が報告されている(Garrido et al.)。
<渡り>
渡り性の種である。ユーラシア大陸の個体群は、サハラ砂漠以南のアフリカとインド亜大陸で越冬するために渡りを行い、9~10月頃に多数が渡る。8月下旬に北方の生息域を離れ、通常10月中旬には越冬地に到着する。3月から4月初旬にかけて春の渡りが見られる。この種は部分的に広い前線上を横断すると考えられているが(地中海全域の島々で定期的に見られることから推測)、それでも多くの渡り鳥が毎シーズン、ボトルネックとなる短い横断地点であるジブラルタル海峡などを通過する。鳥は単独かペアで見られることが多く、渡りのときでも5羽以上の群れを作ることはまれで、他の猛禽類には近づかない(Ferguson-Lees and Christie 2001)。




















































.jpg)



